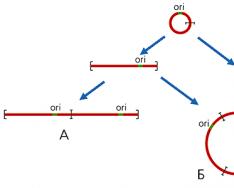
Функционирование митохондриального геномаЧто же особенного в механизмах репликации и транскрипции ДНК митохондрий млекопитающих? У большинства животных комплементарные цепи в мтДНК значительно различаются по удельной плотности, поскольку содержат неодинаковое количе-ство “тяжелых” пуриновых и “легких” пиримидиновых нуклеотидов. Так они и называются - H (heavy - тяжелая) и L (light - легкая) цепь. В начале репли-кации молекулы мтДНК образуется так называемая D-петля (от англ. Displace-ment loop - петля смещения). Эта структура, видимая в электронный микро-скоп, состоит из двуцепочечного и одноцепочечного (отодвинутой части Н-цепи) участков. Двуцепочечный участок формируется частью L-цепи и компле-ментарным ей вновь синтезированным фрагментом ДНК длиной 450-650 (в зависимости от вида организма) нуклеотидов, имеющим на 5"-конце рибонук-леотидную затравку, которая соответствует точке начала синтеза Н-цепи (oriH). Синтез L-цепи начинается лишь тогда, когда дочерняя Н-цепь доходит до точки ori L. Это обусловлено тем, что область инициации репликации L-цепи доступ-на для ферментов синтеза ДНК лишь в одноцепочечном состоянии, а следовате-льно, только в расплетенной двойной спирали при синтезе Н-цепи. Таким обра-зом, дочерние цепи мтДНК синтезируются непрерывно и асинхронно (рис.3). Рис 3. В митохондриях общее число молекул с D-петлей значительно превыша-ет число полностью реплицирующихся молекул. Обусловлено это тем, что у D-петли есть дополнительные функции - прикрепление мтДНК к внутренней ме-мбране и инициация транскрипции, поскольку в этом районе локализованы промоторы транскрипции обеих цепей ДНК. В отличие от большинства эв-кариотических генов, которые транскрибируются независимо друг от друга, ка-ждая из цепей мтДНК млекопитающих переписывается с образованием одной молекулы РНК, начинающейся в районе ori H. Помимо этих двух длинных мо-лекул РНК, комплементарных Н- и L-цепям, формируются и более короткие участки Н-цепи, которые начинаются в той же точке и заканчиваются на 3"-кон-це гена 16S рРНК (рис.4). Таких коротких транскриптов в 10 раз больше, чем длинных. В результате созревания (процессинга) из них образуются 12S рРНК и 16S рРНК, участвующие в формировании митохондриальных рибосом, а так-же фенилаланиновая и валиновая тРНК. Из длинных транскриптов вырезаются остальные тРНК и образуются транслируемые мРНК, к 3"-концам которых при-соединяются полиадениловые последовательности. 5"-концы этих мРНК не кэ-пируются, что необычно для эвкариот. Сплайсинга (сращивания) не происхо-дит, поскольку ни один из митохондриальных генов млекопитающих не содер-жит интронов. |
|
 ND1-ND6, ND4L - гены субъединиц НAД-H-дегидрогеназного комплекса; СОI-COIII - гены субъединиц цитохром-с-оксидазы; ATP6, ATP8 - гены субъединиц AТФ-синтетазы Cyt b - ген цитохрома b. |
|
Рис 4. Транскрипция мтДНК человека, содержащей 37 генов. Все транскрипты начинают синтезироваться в районе ori H. Рибосомные РНК вырезаются из длинного и короткого транскриптов Н-цепи. тРНК и мРНК образуются в результате процессинга из транскриптов обеих цепей ДНК. Гены тРНК обозначены светло-зеленым цветом.
Хотите узнать какие еще сюрпризы способен преподнести митохон-дриальный геном? Отлично! Читаем дальше!..
Несмотря на то, что в геномах митохондрий млекопитающих и дрожжей содержится приблизительно одинаковое количество генов, размеры дрожжево-го генома в 4-5 раз больше - около 80 тыс. пар нуклеотидов. Хотя кодирую-щие последовательности мтДНК дрожжей высоко гомологичны соответствую-щим последовательностям у человека, дрожжевые мРНК дополнительно имеют 5"-лидерную и 3"-некодирующую области, как и большинство ядерных мРНК. Ряд генов содержит еще и интроны. Так, в гене box, кодирующем цитохром-оксидазу b, имеется два интрона. Из первичного РНК-транскрипта автокатали-тически (без участия каких-либо белков) вырезается копия большей части пер-вого интрона. Оставшаяся РНК служит матрицей для образования фермента ма-туразы, участвующей в сплайсинге. Часть ее аминокислотной последовательно-сти закодирована в оставшихся копиях интронов. Матураза вырезает их, разру-шая свою собственную мРНК, копии экзонов сшиваются, и образуется мРНК для цитохромоксидазы b (рис.5). Открытие такого феномена заставило пере-смотреть представление об интронах, как о “ничего не кодирующих последова-тельностях”.

Рис 5.
При изучении экспрессии митохон-дриальных генов Trypanosoma brucei обнаружилось удивительное отклонение от одной из основных аксиом молекулярной биологии, гласящей, что после-довательность нуклеотидов в мРНК в точности соответствует таковой в коди-рующих участках ДНК. Оказалось, мРНК одной из субъединиц цитохром-с-оксидазы редактируется, т.е. после транскрипции изменяется ее первичная структура - вставляется четыре урацила. В результате образуется новая мРНК, служащая матрицей для синтеза дополнительной субъединицы фермента, пос-ледовательность аминокислот в которой не имеет ничего общего с последова-тельностью, кодируемой нередактированной мРНК (см. таблицу).

Наибольший сюрприз ученым митохондрии преподнесли в 1979 г. До то-го времени считалось, что генетический код универсален и одни и те же трип-леты кодируют одинаковые аминокислоты у бактерий, вирусов, грибов, расте-ний и животных. Английский исследователь Беррел сопоставил структуру од-ного из митохондриальных генов теленка с последовательностью аминокислот в кодируемой этим геном субъединице цитохромоксидазы. Оказалось, что гене-тический код митохондрий крупного рогатого скота (как и человека) не просто отличается от универсального, он “идеален”, т.е. подчиняется следующему пра-вилу: “если два кодона имеют два одинаковых нуклеотида, а третьи нуклеоти-ды принадлежат к одному классу (пуриновых - А, Г, или пиримидиновых - У, Ц), то они кодируют одну и ту же аминокислоту”. В универсальном коде есть два исключения из этого правила: триплет АУА кодирует изолейцин, а кодон АУГ - метионин, в то время как в идеальном коде митохондрий оба эти трип-лета кодируют метионин; триплет УГГ кодирует лишь триптофан, а триплет УГА - стоп-кодон. В универсальном коде оба отклонения касаются прин-ципиальных моментов синтеза белка: кодон АУГ - инициирующий, а стоп-кодон УГА останавливает синтез полипептида. Идеальный код присущ не всем описанным митохондриям, но ни у одной из них нет универсального кода. Мож-но сказать, что митохондрии говорят на разных языках, но никогда - на языке ядра.
Различия между “универсальным” генетическим кодом и двумя митохондриальными кодами
Митохондриальный
код млекопитающих
Митохондриальный
код дрожжей
“Универсальный”
ДНК в митохондриях представлена циклическими молекулами, не образующими связь с гистонами, в этом отношении они напоминают бактериальные хромосомы.
У человека митохондриальная ДНК содержит 16,5 тыс. н.п., она полностью расшифрована. Найдено, что митохондральная ДНК различных объектов очень однородна, отличие их заключается лишь в величине интронов и нетранскрибируемых участков. Все митохондриальные ДНК представлены множественными копиями, собранными в группы, кластеры. Так в одной митохондрии печени крысы может содержаться от 1 до 50 циклических молекул ДНК. Общее же количество митохондриальной ДНК на клетку составляет около одного процента. Синтез митохондриальных ДНК не связан с синтезом ДНК в ядре. Так же как и у бактерий митохондральная ДНК собрана в отдельную зону – нуклеоид, его размер составляет около 0, 4 мкм в диаметре. В длинных митохондриях может быть от 1 до 10 нуклеоидов. При делении длинной митохондрии от нее отделяется участок, содержащий нуклеоид (сходство с бинарным делением бактерий). Количество ДНК в отдельных нуклеоидах митохондрий может колебаться в 10 раз в зависимости от типа клеток. При слиянии митохондрий может происходить обмен их внутренними компонентами.
рРНК и рибосомы митохондрий резко отличны от таковых в цитоплазме. Если в цитоплазме обнаруживаются 80s рибосомы, то рибосомы митохондрий растительных клеток принадлежат к 70s рибосомам (состоят из 30s и 50s субъединиц, содержат 16s и 23s РНК, характерные для прокариотических клеток), а в митохондриях клеток животных обнаружены более мелкие рибосомы (около 50s). В митоплазме на рибосомах идет синтез белков. Он прекращается, в отличие от синтеза на цитоплазматических рибосомах, при действии антибиотика хлорамфеникола, подавляющего синтез белка у бактерий.
На митохондриальном геноме синтезируются и транспортные РНК, всего синтезируется 22 тРНК. Триплетный код митохондриальной синтетической системы отличен от такового, используемого в гиалоплазме. Несмотря на наличие казалось бы всех компонентов, необходимых для синтеза белков, небольшие молекулы митохондриальной ДНК не могут кодировать все митохондриальные белки, только лишь их небольшую часть. Так ДНК размером 15 тыс.н.п. может кодировать белки с суммарным молекулярным весом около 6х105. В это же время суммарный молекулярный вес белков частицы полного дыхательного ансамбля митохондрии достигает величины около 2х106.
Рис. Относительные размеры митохондрий у различных организмов.
Интересны наблюдения за судьбой митохондрий в дрожжевых клетках. В аэробных условиях дрожжевые клетки имеют типичные митохондрии с четко выраженными кристами. При переносе клеток в анаэробные условия (например, при их пересеве или при перемещении в атмосферу азота) типичные митохондрии в их цитоплазме не обнаруживаются, и вместо них видны мелкие мембранные пузырьки. Оказалось, что в анаэробных условиях дрожжевые клетки не содержат полную дыхательную цепь (отсутствуют цитохромы b и a). При аэрации культуры наблюдается быстрая индукция биосинтеза дыхательных ферментов, резкое повышение потребления кислорода, а в цитоплазме появляются нормальные митохондрии.
Расселение людей на Земле
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
Генетика митохондрий
1. Формальная генетика митохондрий
В отличие от пластид, митохондрии содержатся у всех эукариот: у растений, животных и грибов. Митохондрии всех трех царств выполняют одну и ту же функцию, и структура у них в целом сходна. Митохондрии представляют собой округлые структуры размером от 1 мкм (рис. 1).
Рис. 1 Электронная микрофотография митохондрий листового мезофилла
Однако, в ряде случаев митохондрии могут быть объединены в достаточно длинную трубчатую изогнутую структуру. Внутреннее содержимое митохондрий называется матрикс. Матрикс содержит тонкие фибриллы и гранулы. Было выяснено, что гранулы -- это митохондриальные рибосомы, отличающиеся по размеру и плотности от рибосом цитоплазмы. Митохондрии, как и другие органеллы, окружены наружной двойной мембраной. Наружная мембрана митохондрий сходна с наружной мембраной пластид, ядра и с мембраной эндоплазматического ретикулума. Внутренняя мембрана митохондрий образует впячивания -- кристы. Именно на поверхности внутренней мембраны расположены все основные ферментные ансамбли, обеспечивающие функции митохондрий. Существуют методы разделения внутренней и внешней мембраны митохондрий. Поскольку наружная мембрана митохондрий менее плотная и необратимо набухает в растворе фосфата, это приводит к ее разрыву и отделению от внутренней. После обработки фосфатом изолированных митохондрий при помощи центрифугирования можно разделить внешние и внутренние мембраны этих органелл. Если посмотреть на них в электронный микроскоп, то они выглядят как прозрачные полые сферы, причем объем сферы, образованной внутренней мембраной, намного выше, чем объем сферы внешней мембраны. Поэтому объемную структуру митохондрий легко представить себе как большой шар, помещенный внутри маленького шара. В этом случае у внутренней мембраны возникнут многочисленные складки, так называемые кристы. Активность процессов, протекающих в митохондриях, непосредственно связана с количеством и размером крист. Чем больше поверхность крист и, следовательно, поверхность внутренней мембраны, тем активнее идут эти процессы. Следовательно, внутренняя мембрана митохондрий изменяется в размерах в зависимости от функционального состояния органелл.
Внутренние и наружные мембраны различаются между собой по плотности (внутренняя более плотная), по проницаемости (внутренняя обладает высокоспецифичной проницаемостью, наружная неспецифичной), разным составом ферментов и разным соотношением белков к липидам.
Внутренняя мембрана митохондрий уникальна по своему строению. Она содержит многокомпонентные комплексы белков-ферментов, которые осуществляют перенос электронов, окислительное фосфолирование, синтез цепи жирных кислот, а также белки, которые регулируют перенос небольших молекул во внутреннюю полость митохондрий.
Митохондрии так же, как и пластиды, никогда не возникают «de novo». Даже у организмов, живущих в анаэробных условиях, структуры, подобные митохондриям, есть. Если, например, выращивать один и тот же штамм дрожжей в аэробных и анаэробных условиях, то в клетках, выросших в анаэробных условиях, изменяется размер митохондрий, но не снижается их количество.
Деление митохондрий так же, как и пластид, осуществляется при помощи амитоза, с образованием гантелевидных фигур и их последующей перешнуровкой.
В некоторых случаях удалось показать синхронность деления митохондрий с ядром клетки и достаточно точное их распределение по дочерним клеткам у некоторых биологических объектов. Так, у инфузорий показана полная синхронность деления митохондрий вместе с ядром клетки. В митотически делящихся клетках растений и делящихся сперматоцитах аскариды было показано, что митохондрии достаточно точно распределяются вдоль веретена деления.
Исторически, почти вся формальная генетика митохондрий была изучена на грибах и, главным образом, на дрожжах. У остальных организмов имеются только отдельные факты связи тех или иных признаков с митохондриями. Жизненный цикл дрожжей представлен на рисунке
Рис. 2 Жизненный цикл Saccharomyces cerevisiae
Дрожжи -- одноклеточный, но многоядерный организм. Значительную часть своей жизни они находятся в гаплофазе и, следовательно, их ядра гаплоидны. Гаплоидные клоны, обладающие противоположными половыми факторами (или типами скрещиваемости), А и а, могут сливаться друг с другом. Гаплоидные клоны с одинаковыми типами скрещиваемости не могут участвовать в оплодотворении. После оплодотворения ядра сливаются и образуются диплоидные клоны. В диплоидных клонах происходит споруляция и мейоз, образуется аск, дающий начало гаплоидным клонам двух противоположных типов скрещиваемости А и а в равных пропорциях. Естественно, что простые менделевские гены будут расщепляться точно так же, как и ген, контролирующий половой фактор, т.е. будут давать расщепление 1:1.
Дрожжи в зиготной фазе являются гетерозиготными и могут размножаться двояко: вегетативным и генеративным путем. При вегетативном размножении они просто делятся, и в образовавшиеся клетки попадает несколько диплоидных ядер. Кроме того, вегетативное размножение может происходить и при помощи почкования. В образовавшихся почках ядра тоже, диплоидные. Естественно, что при вегетативном размножении никакого расщепления ядерных генов не происходит -- гетерозиготы остаются гетерозиготами.
При генеративном размножении происходит мейоз и образуются клетки с гаплоидными ядрами, называемые аскоспорами. Аскоспоры гаплоидны, и расщепление у них происходит на равное количество аскоспор с доминантным и рецессивным аллелями, т.е. 1:1.
Таким образом, если расщепления 1:1 не наблюдается, то это могло бы указывать нам на то, что эти гены, возможно, неменделевские и, следовательно, возможно, цитоплазматические.
Существование внеядерного мутанта у дрожжей впервые продемонстрировал французский исследователь Б. Эффрусси еще в 1949 г. Эти мутанты проявляли дефекты дыхания и слабый рост. Они не содержали некоторые цитохромы. Такие мутанты можно было получить в большом количестве (иногда до 100 %) под действием акридиновых красителей. Но и спонтанно они могут возникать с частотой до 1 %. Эти мутанты получили название «petite », от французского слова «маленький».
При скрещивании этих мутантов с нормальными штаммами все потомство было без исключения нормальным. Хотя по другим генетическим маркерам, таким, как потребность в аденине, тиамине, расщепление по факторам полового типа было нормальным -- 1:1.
Если из первого поколения гибридов случайно выбирать клетки и скрещивать их опять с мутантами petite , все потомство было опять нормальным, правда, иногда появлялись и редкие мутантные выщепенцы с частотой менее 1%. Т.е. они появлялись практически с той же частотой, что и спонтанное возникновение этих мутантов. Можно было опять отбирать эти гибриды и скрещивать с нормальными с тем же результатом. Если исходить из того, что это мутации ядерных генов, то это можно было бы представить, как результат расщепления по 20 независимым локусам. Возникновение мутанта с одновременной мутацией в 20 локусах событие практически невероятное.
Р. Райт и Д. Ледерберг получили убедительные доказательства того, что данные мутанты не являются ядерными. Схема их эксперимента была следующей. При слиянии клеток дрожжей ядра сливаются не сразу, и в этот момент можно отсадить почки, содержащие еще гаплоидные ядра как одного, так и другого родителя. Такие гаплоидные почки спонтанно диплоидизируются (А --> АА; а --> аа). Если один штамм, например, с мутацией petite помечен неспособностью расти на аргинине, а второй -- не petite , помечен неспособностью расти на триптофане, то, отбирая почки у таких гибридов, мы отбираем родительские штаммы по ядерным генам. Что же происходит с цитоплазматическими? В результате эксперимента Р. Райта и Д. Ледерберга было выявлено следующее. Из 91 клона было обнаружено 6 клонов, у которых ядро было такое же, как и у не petite мутанта, а фенотип типичный petite . Следовательно, данный фенотип определяется не ядром, а независимо от него, и данную мутацию можно было назвать неядерной.
В дальнейшем были обнаружены и ядерные мутации petite . Всего таких мутантов было обнаружено около 20. Все они нормально менделировали и потомство аскоспор давало нормальное расщепление 2:2, хотя фенотипически они были очень схожи с цитоплазматическими мутантами. При скрещивании цитоплазматических petite с ядерными было обнаружено, что у зигот появляется способность к нормальному дыханию, а затем происходит расщепление 2: Таким образом, тест на комплементарность доказывал то, что мы имеем дело с мутантами разной локализации. Обнаружение ядерных и цитоплазматических мутантов с нарушенной функцией митохондрий указывало также и на то, что не все функции этих органелл кодируются цитоплазматическими генами. Часть из них кодируют ядерные гены.
В дальнейшем Б. Эффрусси обнаружил еще один такой же фенотип, как petite , но наследование данной мутации происходило другим образом. При скрещивании мутантов petite с нормальными клетками все потомство приобретало свойство медленно расти, а расщепление было 0:4. Первый тип цитоплазматических мутантов, который давал только нормальное потомство, в связи с этим был назван нейтральным, а второй, который давал только мутантное, был назван супрессивным, или доминантным, petite . Супрессивность в данном случае это своего рода доминирование. Но это доминирование особого рода, когда рецессивный аллель не просто скрывается в гетерозиготе, он попросту исчезает совсем. Многочисленные опыты показали, что и супрессивные мутанты petite также являются цитоплазматическими, так как факторы, обуславливающие их появление, не наследуются вместе с ядром.
В дальнейшем молекулярные исследования выявили, что супрессивные мутанты petite в отличие от нейтральных обладают более короткими молекулами митохондриальной ДНК, состоящими практически только из АТ-пар. Вероятнее всего, эффект супрессивности основан на более быстром размножении таких митохондриальных ДНК и вследствие этого вытеснением нормальных митохондриальных ДНК.
Таким образом, в цитоплазматических мутантах типа petite имеют место либо сравнительно небольшие делеции в митохондриальной ДНК (нейтральные мутанты petite ), либо тотальные перестройки митохондриального генома -- (супрессивные мутанты petite ).
Кроме того, были обнаружены мутанты с неполной супрессивностью, т.е. способностью давать определенный процент особей нормального типа 10, 20, 30 и даже около 50 процентов.
Оказалось, что степень супрессивности зависит от воздействий внешней среды -- температуры, субстрата и т.д. Ядерные мутанты не проявляли такой зависимости, что позволило отличать неполно супрессивные цитоплазматические petite от ядерных.
После получения данных о цитоплазматических мутантах устойчивости к антибиотикам у хламидомонады, стали получать мутации устойчивости к антибиотикам и у дрожжей. Целый ряд таких мутантов - также оказался цитоплазматическим. При скрещивании, например, эритромицинчувствительных с эритромицинустойчивыми ERs х ERr , все потомство было эритромицинчувствительным Ers (т.е. таким же, как и дикий тип) и не происходило никакого расщепления. Такой же результат был продемонстрирован и с мутантами устойчивости к другим антибиотикам. Однако, если сразу после образования зиготы отбирать почки, то среди них можно найти и мутантные фенотипы.
При дигибридном скрещивании, т.е. при скрещивании двух цитоплазматических мутантов, чувствительных к разным антибиотикам, например устойчивых к хлорамфениколу, но чувствительных к эритромицину с чувствительными к хлорамфениколу, но устойчивыми к эритромицину CrERs х CsERr , в потомстве преобладал фенотип только одного из родителей -- CrERs . Вместе с тем, при отборе из почек сразу после оплодотворения были обнаружены не только родительские классы фенотипов, но и рекомбинанты: CrERr и CsERs , т.е. чувствительные или устойчивые к обоим антибиотикам. Наличие рекомбинантов впервые показало, что и митохондриальные гены могут рекомбинировать так же, как и ядерные. Вместе с тем, в отличие от экспериментов по рекомбинации пластидных генов у хламидомонады, у дрожжей была обнаружена полярность рекомбинации, т.е. неравное количество рекомбинантных фенотипов в зависимости от направления скрещивания. Полярность рекомбинации была объяснена как наличие специального генетического фактора пола в геноме митохондрий. Этот фактор был обозначен как щ+ и щ-. Родительская форма, имеющий фактор щ+, т.е. женский родитель, обеспечивает преимущественную передачу (более высокую частоту передачи) своих маркеров. При скрещивании однополых по этому митохондриальному фактору родителей полярности рекомбинации не наблюдается и получается равное число рекомбинантов. Сам половой фактор митохондрий наследуется независимо от пола организма.
В действительности имеется ли пол у органелл цитоплазмы -- митохондрий в общепринятом смысле? Можно считать, что имеется, если мы считаем, что у кишечной палочки он есть.
Но главное заключалось в том, что при помощи множества полученных мутаций и обнаружения рекомбинации митохондриальных генов стало возможным их картирование.
В экспериментах по скрещиванию мутаций типа petite с мутациями устойчивости к антибиотикам было выяснено, что, по крайней мере, все супрессивные мутации petite в скрещиваниях утрачивают гены устойчивости к антибиотикам. Как было выяснено, это происходит потому, что супрессивные petite имеют обширные области поражения митохондриальной ДНК, и в этом случае ожидать рекомбинации просто невозможно. При индукции мутаций дыхательной недостаточности у мутантов с устойчивостью к тем или иным антибиотикам оказывалось, что иногда маркеры устойчивости утрачивались. При получении мутантов с дыхательной недостаточностью, используя в качестве исходной формы мутантов с двойной устойчивостью к антибиотикам, у полученных дефектных по дыханию мутантов могли быть потеряны оба маркера устойчивости или только один из них. Это свидетельствовало о том, что мутанты с дыхательной недостаточностью представляют собой ту или иную степень делетирования митохондриальной ДНК, и, следовательно, это также можно было использовать для картирования генома митохондрий.
У нейроспоры в 1952 г. К. Митчелом был обнаружен первый медленно растущий мутант, названный впоследствии MI -1 (сокращение от английского «материнское наследование» -- maternal inheritance ). Наследование этой мутации происходило в зависимости от направления скрещивания, и все потомство было таким же по фенотипу, как и материнская форма. Вероятно, это происходит потому, что при оплодотворении мужская гамета у нейроспоры не привносит цитоплазмы. На связь этой спонтанно возникшей мутации с митохондриями указывало не только материнское наследование и различия в реципрокных скрещиваниях, но и то, что у них отсутствовали цитохромы a и b в системе переноса электронов.
Впоследствии были получены и другие медленно растущие штаммы у нейроспоры, связанные с дыхательной недостаточностью митохондрий. Некоторые из них, например, мутанты MI -3 и MI -4, как оказалось, наследовались так же, как и мутант MI -1, в то время как другая часть, например, мутанты С115 и С117 проявляли обычное менделевское моногибридное наследование. Это напоминает и о других аналогичных случаях, когда фенотип органелл, и хлоропластов, и митохондрий изменяется при возникновении как ядерных, так и цитоплазматических мутаций, что свидетельствует, что и цитоплазматические и ядерные генетические системы совместно контролируют их функции.
В дальнейшем было обнаружено несколько генов супрессоров, введение которых восстанавливало скорость роста у медленно растущих мутантов. Интересно отметить, что каждый из этих супрессоров восстанавливал скорость роста только у одного из мутантов. Например, ген супрессор, названный f , восстанавливал скорость роста у цитоплазматичкого мутанта MI -1, но не у другого цитоплазматического мутанта MI -3 или MI -4, и не у ядерных мутантов С115 и С117 . Аналогичным образом действовали и другие супрессоры. Если через множество поколений путем скрещиваний вывести гены супрессоры из грибов, то мутантный цитоплазматический фенотип снова проявится. Аналогичное взаимодействие ядерных и цитоплазматических генов можно наблюдать и у высших растений, например, при наследовании признака мужской стерильности у многих растений.
При скрещивании ядерных и цитоплазматических медленно растущих мутантов между собой было показано независимое наследование ядерных и цитоплазматических генов.
Например, при скрещивании дикий тип х (MI -1 х С115 ) потомство F 1 (MI -1 х С115 ) было фенотипически однородным -- все особи были медленно растущими, а потомство возвратных или анализирующих скрещиваний дикий тип х (MI -1 х С115 ) уже не содержало мутаций MI -1 и расщеплялось по ядерному гену С-115 в соотношении 1:1.
Скрещивание цитоплазматических мутантов между собой не давало каких-то новых результатов, так как цитоплазматические мутанты, по крайней мере, у нейроспоры при половом воспроизведении демонстрируют строго материнское наследование. Между тем разные цитоплазматические мутанты, хотя и обладали в принципе одинаковым фенотипом -- замедленным ростом -- фенотипические различия между ними все же можно было выявлять, так как они обладали разной степенью замедления этого роста. Однако строгое материнское наследование при половом воспроизведении не позволяло объединить в цитогету (цитоплазматическую гетерозиготу) две цитоплазматические мутации, что делало невозможным рекомбинации цитоплазматических генов и, следовательно, их картирование.
Выход из этого положения был найден при помощи слияния гифов нейроспоры, что позволило соединять в одной клетке различные как ядерные, так и неядерные геномы.
При создании различных цитогет были получены следующие результаты:
MI -1 / дикий тип -- все потомство только дикого типа;
MI -3 / дикий тип -- часть потомства дикого типа, а другая часть растет со скоростью, свойственной мутанту MI -3;
MI -1 / MI -З -- большая часть потомства с фенотипом MI -3 и небольшая часть потомства с фенотипом MI -1;
MI -1 / MI -4 -- первоначально фенотип, свойственный дикому типу, а затем расщепление на фенотипы MI -1 и MI -4.
Таким образом, в последнем случае была обнаружена комплементация цитоплазматических мутаций, что свидетельствует, что эти мутации произошли в разных участках митохондриального генома.
В дальнейшем были получены и другие цитоплазматические мутации нейроспоры. Метод слияния гиф и получения при этом цитогет позволял надеяться на получение различных рекомбинантов и последующего построения генетической карты нейроспоры. Однако этому воспрепятствовало то обстоятельство, что у нейроспоры не было получено большого разнообразия цитоплазматических мутаций такого, как у хламидомонады или дрожжей.
Впоследствии различные нехромосомные мутации, полученные у нейроспоры, исследовали при помощи методов молекулярной биологии, и их удалось связать с митохондриальным геномом.
У другого гриба подоспоры была обнаружена мутация, вызывающая феномен преждевременного старения. У мутантов постепенно снижалась жизнеспособность культуры при пересеве. При реципрокных скрещиваниях был выяснен материнский характер наследования феномена старения. Вместе с тем, материнское наследование было неполным. Передача признака осуществляется как половым путем, так и путем соединения мицелиев. Наличие расщепления, хотя и нерегулярного, указывает на корпускулярную природу наследования признака. Было проведено достаточно много исследований, которые позволили показать, что это неинфекционный агент, а митохондриальный ген. Хотя в настоящее время нет полных молекулярных данных, уже ясно, что это также мутации митохондриального генома. Наличие гена старения в митохондриальном геноме породило массу спекуляций на геронтологические темы, и некоторые медики считают, что старение и у человека связано не только с изменением функций митохондрий, но и с изменением их генома.
Несмотря на спекулятивность идеи о связи геронтологических процессов у человека с изменениями митохондриальной ДНК, новые данные по изучению изменчивости генома митохондрий человека это подтверждают.
Издавна у человека было известно достаточно большое количество заболеваний, наследующихся по материнской линии -- от матери ко всем потомкам. Эти болезни достаточно редко распространены, вероятно, и в силу того, что передаются только женским полом. Кроме того, большие делеционные изменения в митохондриальной ДНК, конечно, чаще всего приводят либо к летальному исходу еще в эмбриональном периоде, либо к нарушению репродукционных функций. В любом случае они эффективно отметаются естественным отбором.
Формальный генетический подход, который был достаточно хорошо применен для исследования цитоплазматических генов у модельных объектов (хламидомонада, дрожжи и др.), не был столь успешным для анализа цитоплазматически наследуемых признаков у человека и поэтому самое большее, что удалось выяснить из анализа родословных, это то, что такие наследственные болезни все же существуют.
Кроме хорошо известного синдрома -- атрофии зрительного нерва (болезнь Лебера или наследственная оптическая невропатия) существуют и другие болезни, наследующиеся по внеядерному типу. Эти болезни связаны, прежде всего, с нарушением функционирования мышц, работы мозга, сердца, эндокринных систем и связаны с недостаточно активной функцией митохондрий в тех или других органах. Существует даже митохондриально обусловленная форма диабета.
Только с помощью молекулярных методов удалось выявить природу этих болезней. Исследование различных семей с болезнью Лебера показало, что в разных случаях имеют место мутации в различных участках митохондриального генома.
Чаще всего семьи с наследственными цитоплазматическими болезнями проявляют гетероплазмию и матери имеют как нормальные, так и мутантные митохондриальные ДНК, в результате чего выщепляются потомки как с мутантным, так и нормальным плазматипом.
Связь между возрастом человека и митохондриальной ДНК также была показана при помощи методов молекулярной биологии. Исследования митохондриальной ДНК у людей различного возраста показали, что у пожилых людей быстро увеличивается процент мутантной митохондриальной ДНК в клетках мозга и сердца. Кроме того, исследования некоторых наследственных синдромов показывают, что обладающие ими пациенты имеют и повышенную частоту мутаций митохондриальной ДНК, что возможно и является причиной сокращения продолжительности жизни.
Кроме мутаций митохондриального генома, приводящих к серьезным патологиям организма, было обнаружено множество, в достаточной степени, нейтральных мутаций митохондриального генома среди различных популяций рас человека. Эти обширные исследования тысяч людей со всех континентов помогают восстанавливать происхождение и эволюцию человека. Сравнивая митохондриальную ДНК человека с ДНК человекообразных обезьян (горилла, орангутанг, шимпанзе) и исходя из того, что дивергенция человека и человекообразных приматов произошла примерно 13 миллионов лет назад, можно рассчитать количество лет, необходимых для изменения одной пары оснований. В дальнейшем, сравнивая дивергенцию митохондриальной ДНК у различных человеческих рас, можно было определить место рождения первой женщины, можно сказать Евы, и время расселения человека по различным континентам (рис. 3).
Размещено на http://www.allbest.ru/
Рис. 3 Расселение человека, согласно данным Д. Уоллеса, по анализу изменчивости митохондриальных ДНК. Цифрами обозначено время заселения данной территории в тысячах лет тому назад
Поскольку самая изменчивая митохондриальная ДНК была обнаружены среди аборигенов Африки, то можно предположить, что «праматерью» человеческого рода была африканская женщина. Произошло это приблизительно 100 000 лет тому назад. Приблизительно 70 000 лет тому назад человек начал заселять центральную Азию через Ближний Восток и Саудовскую Аравию, чуть позже Юго-Восточную Азию, Индонезию и Австралию. Около 50 000 лет тому назад люди появились в Европе. Эти же данные показали, что заселение Американского континента происходило в два этапа: сначала 30 000 лет тому назад через Беренгию (существовавшую в то время сушу, связывающую Америку и Азию) с Севера до самого юга Американского континента, а затем 8 000 лет тому назад также из Северо-Восточной Азии на восток Северной Америки. Переселенцы на островах Тихого океана появились сравнительно недавно -- несколько тысяч лет тому назад.
Следует отметить, что эти данные, основанные на сравнительном анализе митохондриальной ДНК, достаточно хорошо согласуются как с археологическими данными, так и с данными лингвистического анализа.
Использование именно митохондриальной ДНК для анализа истории человечества стало возможным потому, что митохондриальный геном имеет сравнительно небольшой размер, наследуется исключительно по материнской линии и в отличие от ядерных генов не рекомбинирует.
Геном митохондрий
Митохондрии содержатся не только в клетках растений, но также и в клетках животных и грибов. Эти органеллы более универсальны, чем пластиды. Впервые ДНК в митохондриях были открыты в 1963 году (М. Наас) сразу же после открытия ДНК в пластидах. Несмотря на сходство функций и структуры митохондрий всех трех царств эукариот, их генетическая организация достаточно сильно различается, поэтому обычно организацию геномов митохондрий у этих царств рассматривают отдельно, выявляя при этом общие черты организации генома.
Физико-химический состав ДНК митохондрий у различных царств различен. У растений он довольно постоянен: от 45 до 47 % ДНК состоит из ГЦ-пар. У животных и грибов -- варьирует более значительно: от 21 до 50 % ГЦ-пар.
У многоклеточных животных размеры генома митохондрий колеблются от 14.5 до 19.5 т.п.н. Практически, это всегда одна кольцевая молекула ДНК. Например, ДНК митохондрий человека -- кольцевая молекула размером 16 569 пар нуклеотидов. Этот размер можно выразить и в других единицах -- в виде молекулярной массы -- 10 6 дальтон или в виде длины контура молекулы -- 5 мкм. Первичная структура этой молекулы полностью определена. В митохондриях содержится собственный аппарат трансляции -- т.е. собственные рибосомы 70S, похожие на хлоропластные или прокариотические и состоящие из двух субъединиц, собственные матричные РНК, необходимые ферменты и белковые факторы. В их геноме закодированы 12S- и 16S - рибосомальные РНК, а так же 22 транспортные РНК. Кроме того, митохондриальная ДНК кодирует 13 полипептидов, из которых 12 идентифицированы. Все кодирующие последовательности расположены прямо друг за другом. В крайнем случае, они разделены лишь несколькими нуклеотидами. Некодирующие последовательности, т.е. интроны отсутствуют. Вслед за кодирующей последовательностью почти всегда находится ген транспортной РНК. Например, порядок таков: транспортная РНК фенилаланина -- ген 12S рибосомальной РНК -- транспортная РНК валина -- ген 16S рибосомальной РНК -- транспортная РНК лейцина и т.д. Такой порядок характерен не только для митохондрий человека, он очень консервативен и характерен для всех животных: дрозофилы, быка, мыши, птиц, рептилий и др. животных.
Большая часть генов расположена в тяжелой цепи, в легкой цепи только гены восьми транспортных РНК и один структурный ген. Таким образом, в отличие от всех других геномов, в геноме митохондрий обе цепи смысловые.
Хотя порядок генов у митохондрий животных и одинаков, выяснено, что сами гены обладают различной консервативностью. Наиболее вариабельна последовательность нуклеотидов участка начала репликации и ряд структурных генов. Наиболее консервативные последовательности расположены в генах рибосомальных РНК и некоторых структурных генах, в том числе в кодирующей последовательности АТФ-азы.
Следует отметить, что универсальность генетического кода нарушена в геноме митохондрий. Например, митохондрии человека используют триплет AUA в качестве кодона для метионина, а не для изолейцина, как у всех, а триплет UGA, используемый в стандартном генетическом словаре как терминирующий кодон, у митохондрий кодирует триптофан.
В целом митохондриальная ДНК человека выглядит так же, как и других млекопитающих: мыши и быка. Несмотря на то, что это далеко не близкие виды -- размеры их митохондриальных ДНК довольно близки между собой: 16 569; 16 295; и 16 338 пар оснований, соответственно. Гены транспортной РНК разделяют некоторые смысловые гены. Наиболее важные из структурных генов -- гены цитохромоксидазы, NADH-дегидрогеназы, цитохром-С оксидоредуктазы и АТФ-синтетазы (рис. 4).
На карте митохондриального генома человека, кроме генов показано и пять хорошо известных болезней человека, наследующихся по материнской линии и вызванных мутациями в митохондриальном геноме.
Так, например, болезнь Лебера -- атрофия зрительного нерва -- вызвана мутацией в гене NADH дегидрогеназы. Эта же болезнь может быть вызвана и мутацией в гене цитохрома b и других локусов. Всего известно нарушение четырех локусов, способных вызвать тот же мутантный фенотип. Кроме того, на этой же карте показано еще четыре болезни, связанные с дефектами мозга, мышц, сердца, почек и печени. Все эти болезни наследуются по материнской линии, и если мать имеет не только дефектные, но и нормальные митохондриальные ДНК и митохондрии, то, происходит сортировка мутантных и нормальных органелл, и потомство может иметь и те, и другие органеллы в различных пропорциях, и мы можем наблюдать также и соматическое расщепление, когда отдельные части тела не будут иметь этих дефектов.
Размещено на http://www.allbest.ru/
Рис. 4 Структура митохондриального генома млекопитающих, основанная на полном сиквенсе митохондриальной ДНК человека, мыши и быка
Таким образом, небольшой по размерам митохондриальный геном животных может кодировать чрезвычайно важные функции организма и в значительной степени определять его нормальное развитие.
Так же, как и геном пластид, геном митохондрий кодирует только часть митохондриальных полипептидов (табл. 1) и наблюдается феномен двойного кодирования. Например, часть субъединиц АТФ-азного комплекса кодируется ядром, в то время как другая часть -- геномом митохондрий. Большая часть генов, кодирующих рибосомальные миохондриальные РНК и белки, а также ферменты транскрипции и трансляции, кодируется ядром клетки.
Таблица 1
Гены митохондриальных ДНК животных
митохондрия геном нейроспора мезофилл
генома животных:
1. компактное расположение генов на мтДНК;
отсутствие в генах интронов;
3. отсутствие некодирующих участков в мтДНК, кроме областей ORI;
4. расположение генов тРНК между другими генами;
5. высокое сходство в размерах генома и в расположении генов у различных видов;
6. наличие одного ORI для каждой нити мтДНК;
7. симметричная транскрипция обеих нитей;
8. наличие одной, в принципе, области инициации транскрипции для каждой нити ДНК;
9. отсутствие 5 / - и 3 / - концевых некодирующих последовательностей в мРНК;
10. созревание мРНК в результате расщепления первичного транскрипта по последовательностям тРНК.
У грибов размеры генома митохондрий в среднем значительно больше и колеблются от 17,3 до 101 т.п.н. Причем в дополнении к основной, как правило, кольцевой молекуле ДНК обнаруживают и от одной до 4-х плазмидоподобных кольцевых или линейных молекул размером от 1 до 13 т.п.н. Размеры митохондриального генома у дрожжей варьируют не только между различными видами, но и даже между различными штаммами. Основные причины существенных различий в геноме митохондрий у грибов заключаются в наличии - отсутствии интронов. У разных видов дрожжей, например, размер митохондриальной ДНК колеблется от 57 до 85 т.п.н.
Наличие интронов и молекул митохондриальной ДНК различного размерного класса является наиболее характерной чертой, отличающей митохондрии грибов от митохондрий животных. Интроны разрывают многие последовательности -- гены рибосомальной РНК, гены некоторых структурных белков, кодирующих митохондриальные ферменты. Наличие большинства интронов не является обязательным для нормального функционирования митохондрий. Искусственно сконструированы штаммы дрожжей, полностью лишенные митохондриальных интронов.
Многие интроны митохондриальной ДНК дрожжей содержат открытые рамки считывания, которые кодируют мутуразы, участвующие в сплайсинге, тогда как другие интроны содержат кодирующие последовательности эндонуклеаз и даже обратных транскиптаз.
Все гены, обнаруженные в митохондриальной ДНК животных, присутствуют и у грибов. Кроме того, у грибов обнаружены и другие гены: у них большее число генов тРНК, обнаружены гены 6-й, 8-й и 9-й субъединиц АТФ-азного комплекса, ряд новых структурных генов и ряд генов с неизвестной функцией (табл. 2).
Таблица 2
Гены митохондриальных ДНК дрожжей
|
Компоненты митохондрий |
||
|
Рибосомальные РНК |
rns(21 S), rnl(1 5 S) |
|
|
Рибосомальные белки: малая субъединица |
||
|
Транспортные РНК |
||
|
Цитохром b (комплекс III) |
с ob (или cyb ) |
|
|
Цитохром с оксидаза (комплекс IV) |
cox 1, cox 2, coxd 3 |
|
|
АТФ-синтаза |
atp6, atp8, atp9 |
|
|
Интрон-кодируемые off: РНК-матуразы Эндонуклеазы Белки, подобные обратной трансриптазе |
aI1, aI2 |
|
|
Неидентифицированные рамки считывания |
В митохондриальной ДНК дрожжей обнаружено только 2 гена рибосомальной РНК и только 1 ген рибосомальных белков. Этот белок расположен в малой субъединице рибосомы. Ген рибосомального белка достаточно вариабелен по размерам даже у разных штаммов, за что и получил название вариабельного (Var l). Остальные белки и РНК митохондриальных рибосом кодируются ядерными генами. 24 гена транспортной РНК обеспечивают транспорт всех аминокислот к месту синтеза белка и только одна транспортная РНК, транспортирующая лизин, импортируется из цитоплазмы и кодируется ядром. Все транспортные РНК митохондрий дрожжей кодируются одной и той же нитью ДНК и только одна из них -- противоположной нитью. Ни один из генов транспортной ДНК не имеет интронов. Гены белков цитохрома b и гены белков цитохрома С могут иметь множество интронов -- от 5 до 9.
Из приведенных данных следует, что кодируемых митохондриальным геномом дрожжей структурных белков явно недостаточно для функционирования этих органелл и большая часть из них кодируется ядерным геномом.
Характерные черты организации и экспрессии митохондриального генома грибов:
1. значительное разнообразие в наборах и расположении митохондриальных генов у различных видов;
большое разнообразие способов организации генетического материала - от компактной организации генома до свободного распределения генов по мтДНК при протяжённых некодирующих последовательностях между генами;
3. мозаичное строение ряда генов;
4. значительные внутривидовые вариации размеров мтДНК, связанные с наличием "необязательных" интронов;
5. способность отдельных сегментов мтДНК к выщеплению и амплификации с образованием дефектного митохондриального генома;
6. наличие одного или нескольких ORI, в каждом из которых репликация инициируется двунаправлено;
7. расположение всех митохондриальных генов на одной нити мтДНК и асимметричная транскрипция мтДНК;
8.множественность транскрипционных единиц мтДНК;
9. разнообразие сигналов процессинга первичных транскриптов, в качестве которых могут использоваться и тРНК, и олигонуклеотидные блоки другого типа - в зависимости от вида;
10. в большинстве случаев мРНК содержат протяжённые концевые некодирующие последовательности.
Наиболее сложная организация митохондриального генома у высших растений. У них митохондриальный геном представляет собой набор суперскрученных двуцепочечных кольцевых и/или линейных молекул. Все последовательности митохондриального генома могут быть организованы в одну крупную кольцевую «хромосому», а наблюдаемые различные размерные классы митохондриальных ДНК, скорее всего, являются результатом рекомбинационных процессов. По крайней мере, на шпинате, видах двух родов Brassica и Raphanus , сахарной свекле и пшенице было показано, что причина такой диспергированности митохондриального генома заключается в рекомбинации гомологичных участков митохондриальной ДНК. Благодаря наличию прямо ориентированных двух-трех семейств повторов размером от 1 до 14 т.п.н., молекулы митохондриальной ДНК способны к активным меж- и внутригеномным перестройкам. В результате таких перестроек митохондриальная ДНК может присутствовать в виде молекул различного размерного класса.
Так, например, у крестоцветного Brassica campestris митохондриальная ДНК присутствует в виде кольцевых молекул трех типов. Первый тип содержит полный геном -- 218 т.п.н., второй - 135 и третий -- 83 т.п.н. Субгеномные кольца образуются в результате рекомбинации геномных колец, имеющих пару прямых повторов длиной 2 т.п.н.
У пшеницы размер митохондриального генома значительно больше -- 430 т.п.н., и имеется более 10 прямых рекомбинационных повторов, в результате при электронно-микроскопическом наблюдении удается увидеть множество колец различного размера, но никто не наблюдал одну большую кольцевую молекулу, возможно, в этом состоянии митохондриальный геном пшеницы никогда и не присутствует. У мха маршанции и другого крестоцветного Brassica hirta прямые рекомбинационные повторы отсутствуют и, возможно, именно поэтому митохондриальная ДНК находиться в виде кольцевых молекул одного размерного класса. Однако для митохондриальной ДНК высших растений это скорее исключение, чем правило. У большинства высших растений в митохондриальном геноме присутствуют как рекомбинационные повторы, так и молекулы митохондриальной ДНК различного размерного класса.
Количество молекул одного размерного класса может различаться очень значительно в различных тканях растения, в зависимости от состояния растений и условий окружающей среды. Отмечено изменение численных соотношений молекул митохондриальной ДНК разного размерного класса при культивировании растений in vivo и in vitro . Возможно, изменение численных отношений между молекулами различных размерных классов отражает адаптивность растений путем повышенной амплификации нужных генов.
Кроме того, в митохондриальной геноме могут присутствовать и плазмиды как линейные, так и кольцевые, как с ДНК-ми, так и РНК-ми последовательностями, размером от 1 до 30 т.п.н. Митохондриальные плазмиды, вероятно, произошли от других клеточных геномов или даже других организмов. Иногда их наличие или отсутствие удается связать с цитоплазматической мужской стерильностью растений, но, правда, не всегда. Плазмиды у некоторых видов присутствуют, а стерильности не наблюдается. По крайней мере, в одном случае достаточно четко продемонстрировано, что в митохондриях линий с так называемым S-типом стерильности кукурузы обнаружена корреляция между присутствием плазмидоподобной митохондриальной ДНК и проявлением феномена цитоплазматической мужской стерильности. Была отмечена способность митохондриальных плазмид встраиваться, как в геном митохондрий, так и в хромосомы ядра. Однако, в других случаях присутствие плазмидной ДНК не всегда вызывает стерильность пыльцы.
Размер митохондриального генома растений наиболее вариабелен -- от 200 до 2500 т.п.н. Размер митохондриального генома высших растений больше, чем размер их хлоропластного генома.
Значительное варьирование размеров митохондриального генома является второй особенностью митохондриального генома растений. Геном не только очень велик, но и может быть различным, даже у близкородственных видов, причем в одних случаях может наблюдаться низкая вариабельность -- виды рода Brassica , в других -- очень большая. Самая высокая вариабельность размеров наблюдается у тыквенных. В пределах этого семейства размер митохондриального генома наиболее изменчив -- от 330 т.п.н. у арбуза до 2500 т.п.н. у дыни. Поэтому доля митохондриальной ДНК в общем объеме генома растений также может значительно изменяться -- порядка 1 % у большинства растений, до 15 % в клетках гипокотиля дыни.
Наличие у митохондриальных геномов больших размеров пытаются объяснить различными причинами.
Наличие дополнительных генов или особых последовательностей, необходимых для функционирования митохондрий.
Присутствие ДНК, которая используется растением, но не в качестве кодирующей, а какой-то другой функции.
ДНК, которая не используется для функционирования митохондрий, так называемая «эгоистичная» ДНК.
По-видимому, существует еще одна возможность для увеличения размеров митохондриального генома -- это последовательности, гомологичные ядерной и хлоропластной ДНК. Последовательности, гомологичные ядерной ДНК, например, у арабидопсис составляют до 5 % митохондриального генома. Первоначально последовательность хлоропластного генома, инкорпорированная в митохондриальный, была обнаружена у кукурузы. Она включала участок около 14 т.п.н., содержащий измененные хлоропластные гены 16S-рибосомальной РНК и участок большой субъединицы РДФК/О. Впоследствии хлоропластные вставки были обнаружены в митохондриальном геноме многих видов высших растений. Обычно, они составляют 1 -- 2 % от митохондриальных последовательностей и включают три главные последовательности.
Последовательность длиной 12 т.п.о. из обратного повтора хлоропластной ДНК. Она содержит последовательности для 3" экзона четырех транспортных РНК и последовательность 16S рибосомальной РНК.
Последовательность длиной от 1,9 до 2,7 т.п.н., которая полностью кодирует большую субъединицу РБФК/О.
Последовательность длиной не более 2 т.п.о. В хлоропластом геноме этот участок кодирует 3" -- конец 23S-pибоcoмальной РНК, 4,5S- и 5S-pPHK, а также три транспортные РНК. Из всех последовательностей хлоропластного генома, которые присутствуют в митохондриальном геноме растений, только последовательности транспортной РНК действительно транскрибируются.
Поскольку у многих видов растений в митохондриальном геноме присутствуют одни и те же хлоропластные последовательности, можно предположить, что они имеют какое-либо функциональное значение. В то же время их роль, механизм переноса и время этого переноса остаются неизвестными. Произошел ли этот перенос в отдаленное в эволюции время становления эукариотической клетки или же наличие хлоропластных вставок в митохондриальном геноме свидетельствует о том, что это нормальный процесс обмена информации между органеллами, происходящий и сейчас, либо он происходит периодически в относительно недавнем эволюционном времени становления конкретных видов и родов растений?
Кроме того, часть последовательностей митохондриального генома представляет собой последовательности, гомологичные вирусным.
Для установления числа генов в геноме митохондрий растений, которые действительно функционируют, рядом исследователей было определено количество продуктов трансляции. Было показано, что число обнаруживаемых белковых полос было одинаковым даже для растений с 10-кратными различиями по величине генома. Хотя использованные методы и не дают прямого ответа на вопрос об общем числе генов митохондриального генома, тем не менее, интересно, что одно и то же число продуктов трансляции было выявлено у анализируемых видов покрытосеменных и было близко к числу генов, кодирующих белки у митохондрий животных и дрожжей.
Впервые полная нуклеотидная последовательность митохондриальной ДНК у растений была определена в 1986 г. у одного вида -- маршанции (Marchantia polymorpha ), а позже у арабидопсиса и у нескольких видов водорослей.
Молекула митохондриальной ДНК у маршанции имеет размер 186 608 п.н. Она кодирует гены 3 рРНК, 29 генов для 27 тРНК и 30 генов известных функциональных белков (16 рибосомальных белков, 3 субъединицы цитохром С оксидазы, цитохрома b, 4 субъединицы АТФ-синтетазы и 9 субъединиц НАДН-дегидрогеназы). Геном также содержит 32 неидентифицированные открытые рамки считывания. Кроме того, обнаружено 32 интрона, расположенных в 16 генах. У разных растений количество генов того или иного комплекса может варьировать, поскольку один или более генов этого комплекса могут быть перенесены в ядро. Среди неидентифицированных генов, по крайней мере, 10 постоянно встречаются практически у всех видов растений, что свидетельствует о важности их функций.
Число митохондриальных генов, кодирующих транспортные РНК митохондрий растений, в значительной степени изменчиво. У многих видов собственных митохондриальных транспортных РНК явно недостаточно, и поэтому они экспортируются из цитоплазмы (кодируются ядром или пластидным геномом). Так, например, у арабидопсиса 12 транспортных РНК имеют митохондриальное кодирование, 6 -- хлоропластное и 13 -- ядерное; у маршанции 29 -- митохондриальное и 2 -- ядерное, и ни одна из транспортных РНК не имеет хлоропластного кодирования; у картофеля 25 -- митохондриальное, 5 -- хлоропластное и 11 -- ядерное; у пшеницы, 9 -- митохондриальное, 6 -- хлоропластное и 3 -- ядерное (табл. 3).
В отличие от митохондриальной ДНК животных и хлоропластных генов, гены митохондриальной ДНК растений дисперсно распределены в геноме. Это касается как генов, кодирующих транспортные РНК, так и генов, кодирующих белки.
Таблица 3
Природа митохондриальных транспортных РНК у растений
|
Количество транспортных РНК, кодируемых геномами |
||||
|
органелл |
||||
|
митохондрий |
хлоропластов |
|||
|
Арабидопсис |
||||
|
Маршанция |
||||
|
Картофель |
||||
|
Не определено |
Не определено |
|||
|
Подсолнечник |
Не определено |
|||
|
Не определено |
||||
|
Кукуруза |
Не определено |
Как и геном митохондрий грибов, геном митохондрий растений имеет интроны, которых не имеют геномы митохондрий животных.
У некоторых видов ряд генов в геноме продублирован. Так, у кукурузы и кормовых бобов гены рРНК не повторены, а у пшеницы они повторены несколько раз. Гены, кодирующие белки митохондрий, также могут быть повторены в их геноме.
Естественно, что митохондрии так же, как и хлоропласты, содержат гораздо больше белков-ферментов, чем их геном генов. И, следовательно, большинство белков контролируются ядерным геномом, собираются в цитоплазме на цитоплазматических, а не на митохондриальных рибосомах и транспортируются в митохондриальные мембраны.
Таким образом, митохондриальный геном растений представляет собой крайне изменчивую по структуре, но достаточно стабильную по числу генов систему. В отличие от компактного генома хлоропластов, в митохондриальном геноме растений гены составляют менее 20 % генома. Увеличение генома митохондрий по сравнению с грибами или животными вызвано наличием интронов, различных повторяющихся последовательностей, вставок из генома хлоропластов, ядра и вирусов. Функции примерно 50 % митохондриального генома растений пока не выяснены. Кроме того, что многие структурные гены, контролирующие функцию митохондрий, находятся в ядре, там же находятся и многие гены, контролирующие процессы транскрипции, процессинга, трансляции митохондриальных генов. Следовательно, митохондрии являются даже менее автономными органеллами, чем пластиды.
Литература
Основная:
1. Алехина Н.Д., Балнокин Ю.В., Гавриленко В.Ф. и др. Физиология растений. Учебник для студ. Вузов. М.: Академия. 2005. 640 с.
Давыденко О.Г. Нехромосомная наследственность. Минск: БГУ. 2001. 189 с.
3. Даниленко Н.Г., Давыденко О.Г. Миры геномов органелл. Минск: Техналогия. 2003. 494 с.
4. Иванов В.И. и др. Генетика. М.: Академкнига. 2006. 638 с.
5. Жимулев И.С. Общая и молекулярная генетика. Новосибирск: Сиб. унив. 2007. 479 с.
6. Сингер М., Берг П. Гены и геномы. М.: Мир. 1998. Т. 1-
7. Ченцов Ю. С. Введение в клеточную биологию. М.: Академкнига. 2004. 495 с.
Дополнительная:
1. Даниленко Н.Г. РНК-редактирование: генетическая информация корректируется после транскрипции // Генетика. 2001. Т. 37. №3. С. 294-316.
Маргелис Л. Роль симбиоза в эволюции клетки. М.: Мир, 1983.
3. Одинцова М. С., Юрина Н. П. Геном митохондрий протистов // Генетика. 200 Т. 38. №6. С. 773-778.
4. Одинцова М. С., Юрина Н. П. Геном пластид высших растений и водорослей: структура и функции // Мол. Биол. 2003. Т. 37. № 5. С. 768-783.
5. Юрина Н. П., Одинцова М. С. Общие черты организации генома хлоропластов. Сравнение с геномами про- и эукариот // Мол. Биол. 199 Т. 36. № 4. С. 757-771.
6. Юрина Н. П., Одинцова М. С. Сравнительная характеристика структурной организации геномов хлоропластов и митохондрий растений// Генетика. 1998. Т. 34. №1. С. 5-2.
Размещено на Allbest.ru
...Подобные документы
Сущность ультраструктурной организации митохондрий. Роль митохондрий в поддержании окислительно-восстановительного баланса клетки. Специфика энергетических функций митохондрий. Изменение морфофункциональных характеристик митохондрий при ацидозе.
дипломная работа , добавлен 27.01.2018
Исследование функциональной роли и структурной организации митохондрий. Рассмотрение и характеристика работы дыхательной цепи митохондрий в условиях нормоксии. Ознакомление с антигипоксическим действием нейротрофического фактора головного мозга.
курсовая работа , добавлен 18.04.2018
Основные механизмы клеточной гибели. Митохондрия как центральный контрольный пункт апоптоза. Морфологические изменения и перераспределение митохондрий в клетке во время апоптоза. Модели высвобождения цитохрома С. Роль митохондрий в процессе старения.
курсовая работа , добавлен 07.01.2013
Комплекс ферментов, локализованных на внутренней мембране митохондрий. Процесс окислительного фосфорилирования. Синтез АТФ на внутренней мембране митохондрий в присутствии кислорода. Компоненты дыхательной цепи. Суть хемиосмотической теории П. Митчелла.
презентация , добавлен 22.10.2014
Изучение плана строения митохондрий и пластид, их функций. Гипотеза о симбиотическом происхождении митохондрий и хлоропластов. Общая типовая характеристика мышечной ткани. Сперматогенез, его основные периоды: размножение, рост, созревание и формирование.
контрольная работа , добавлен 11.03.2014
Понятие и свойства митохондрий, их строение, участие в клеточном дыхании и обмене энергией. Характерные особенности гаструляции эмбрионального развития. Рассмотрение функций, строения, классификации лейкоцитов. Внешний вид тимуса (вилочковой железы).
контрольная работа , добавлен 21.04.2015
Строение, химический состав, распространение в природе и значение таксономической группы слизевиков. Вегетативные тела слизевиков. Трофическая и расселительная стадии. Процесс образования спор. Присутствие в циклах подвижных стадий, строение митохондрий.
курсовая работа , добавлен 12.08.2015
Строение и основные компоненты клеточной оболочки водорослей. Случаи беспорядочного расположения фибрилл среди зеленых водорослей, организация цитоплазмы у различных у различных представителей вида, назначение жгутиков, митохондрий и хлоропластов.
курсовая работа , добавлен 29.07.2009
Клиническое применение фотодинамической терапии. Механизм действия фотосенсибилизаторов на клеточном уровне. Роль митохондрий и ионов кальция в фотодинамически индуцированном апоптозе. Участие сигнальных процессов и защитных белков в реакциях клеток.
контрольная работа , добавлен 19.08.2015
Митохондрия - двумембранная гранулярная или нитевидная органелла, элемент эукариотических клеток (автотрофов и гетеротрофов), энергетическая станция. Основная функция и энергообразование; происхождение, структура. Митохондриальная ДНК и наследственность.
Геном митохондрий человека представлен кольцевой двухцелочечной молекулой ДНК, содержащей 16559 п.н. Доля митохондриальной ДНК от общего количества ДНК достигает 5%. Митохондриальная молекула ДНК состоит из тяжелой (Н) и легкой (L) - цепей. Цепи различаются по нуклеотидному составу. Н-цепь (heavy) содержит больше пурина, легкая L-цепь (ligbt) - больше пиримидина. Митохондриальный геном человека, как и других организмов, представляет собой полуавтономную генетическую систему. Большая часть генов человека локализована в хромосомах ядра, и меньшая - в митохондриальном геноме. 1987г-Адан Уилсон исследовал ДНК 147 представителей различных расс(женщин). Анализ показал, что все мтДНК можно представить как происходящие от одной предковой. Общаа праматерь, к которой восходят все типы мтДНК современных людей, жила в Восточной Африке менее 200тыс лет назад. Митохондрии - это внутриклеточные органеллы, имеющие небольшую собственную хромосому. В отличие от ядерной ДНК, которая содержит подавляющее большинство генов и в процессе полового размножения подвергается рекомбинации, так что потомки получают половину генов от отца, а вторую половину от матери, митохондрии и их ДНК ребёнок получает только из материнской яйцеклетки. Поскольку митохондриальная ДНК не подвергается рекомбинации, изменения в ней могут происходить исключительно посредством редких случайных мутаций. Митохондриа́льные заболева́ния - группа наследственных заболеваний, связанных с дефектами в функционировании митохондрий, приводящими к нарушениям энергетических функций в клетках эукариотов, в частности - человека. Митохондриальные заболевания обусловлены генетическими, структурными, биохимическими дефектами митохондрий, приводящими к нарушениям тканевого дыхания. Они передаются только по женской линии к детям обоих полов, так как сперматозоиды передают зиготе половину ядерного генома, а яйцеклетка поставляет и вторую половину генома, и митохондрии. Патологические нарушения клеточного энергетического обмена могут проявляться в виде дефектов различных звеньев в цикле Кребса, в дыхательной цепи, процессах бета-окисления и т. д. Эффекты митоходриальных заболеваний очень разнообразны. Из-за различного распределения дефектных митохондрий в разных органах, мутация у одного человека может привести к заболеванию печени, а у другого - к заболеванию мозга. Величина проявления дефекта может быть большой или малой, и она может существенно изменяться, медленно нарастая во времени. Некоторые небольшие дефекты приводят лишь к неспособности пациента выдерживать физическую нагрузку, соответствующую его возрасту, и не сопровождаются серьёзными болезненными проявлениями. Другие дефекты могут быть более опасны, приводя к серьёзной патологии.В общем случае, митоходриальные заболевания проявляются сильнее при локализации дефектных митохондрий в мышцах, мозге, нервной ткани, поскольку эти органы требуют больше всего энергии для выполнения соответствующих функций. Для постановки диагноза митохондриального заболевания важен комплексный генеалогический, клинический, биохимический, морфологический и генетический анализ
Presentation Transcript
Синдром Лебера: LHON (1871 г.) наследуемая по материнской линии потеря зрения происходит у людей 20-30 лет вследствие атрофии зрительного нерва и дегенерации ганглиозного слоя клеток ретины Заболевание связано с передаваемой от матери мутацией митохондриальной ДНК в одном из ND генов (комплекс I). В 70% случаев это G11778A(ND4), а в Японии в 90% в 13% случаев G3460A (ND1); в 14% случаевT14484C (ND6) Мутация находится в гомоплазматическом состоянии
634 п.н. ДНК-диагностика синдрома Лебера в семье Nпроведена нами впервые в 2006 году G11778 G11778A замена пробанд с синдромом Лебера здоровый сестра мать человек пробанда
В 80-85% случаев поражаются мужчины (Х хромосома несет какой-то локус чувствительности?) Лишь у 50% мужчин и 10% женщин носителей патогенных мутаций комплекса I в действительности происходит потеря зрения?? Чаще всего мутации, ведущие к синдрому Лебера, встречаются в мтДНК гаплогруппы J; эту группу несут около 15% европейцев?? В формировании заболевания участвуют какие-то дополнительные факторы (???)
Самая часто встречающаяся точечная мутация: А3243Gв лейциновой тРНК Обнаружена у большинства больных с синдромом MELAS инсультоподобные(stroke-like) эпизоды Миопатия лактат-ацидоз энцефалопатия Мутация встречается исключительно в гетероплазматическом состоянии В одних семьях А3243Gвызывает преимущественно кардиомиопатию, в других – диабет и глухоту, в третьих PEO, в четвертых - энцефалопатию???
Синдрома MELAS была проведена нами в 2007 году Мама: фенотипически здоровая женщина очень маленького роста I брак II брак 2ой ребенок 1991-2007 Менингоэнцефалит Умерот ишемического инфаркта обоих полушарий мозжечка 3ий ребенок родился в 1998 Прогрессирующая миопатия, миокардио-дистрофия 1ый ребенок 1988-2000 Кардиопатия, ЗПР, ЗФР. Умерла скоропостижно после травмы Митохондриопатия?? Обнаружена мутация MELAS у сына (80% мутантных молекул в крови) у мамы(40%)
РНК(продолжение) Мутация А8344Gв гене лизиновой тРНК при уровне мутантных молекул > 85% приводит к синдрому MERRF: Миоклонус-эпилепсия; «рваные» красные мышечные волокна; задержка умственного развития; атаксия; атрофия мышц и др. Матери больных обычно фенотипически здоровы или несут слабо выраженные симптомы Мутация резко снижает эффективность трансляции в мт и тем самым провоцирует дефицит дыхательной цепи
Чаще всего встречается мутация гена 12S рРНК A1555G Вызывает несиндромную потерю слуха из-за чувствительности носителей мутации к ототоксическим аминогликозидам Другие мутации генов 12S и 16S вызывают кардиомиопатию, атаксию, MELAS, диабет mellitus, сенсорно-невральная потерю слуха
NARP (neuropathy ataxia and retinitis pigmentosa) Мутация в генеATPase6– трансверсияТ – G в нуклеотиде 8993 (70-90% мутантной ДНК) T8993G:лейцин замещаетсянааргинин вATPase6, чтоприводит к нарушению синтеза АТФ Если доля мтДНК больше 90%, клиническое проявление наблюдается раньшеи симптомы более тяжелые:подострая некротизирующая энцефалопатия с чертами синдрома Лея (LS)
Нейродегенеративное заболевание: - симметричные некротические повреждения в субкортикальных областях ЦНС – базальных ганглиях, таламусе, стволе мозга, спинном мозге; - демиелинизация, сосудистая пролиферация и «глиозис»; - моторная и умственная регрессия, атаксия, дистония, аномальное дыхание Заболевание начинается в раннем детстве, редко во взрослом состоянии; Смерть наступает обычно через два года после начала заболевания
ДНК (MILS) 7/10 cлучаев – рецессивные мутации ядерных аутосомных генов, кодирующих субъединицы дыхательной цепи или белки, участвующие в ее сборке ATPase 6 LS 1/10 cлучаев – мутации Х-хромосомы PDHC
Причина – крупная делеция 5 т.п.н. Утрачиваются 5 генов тРНК и 5 белковых генов KSS –фатальная мультисистемная патология, проявляется в возрасте 4-18 лет:CPEO, пигментный ретинит, атаксия,глухота, эндокринная дисфункция, атриовентрикулярная блокада сердца, повышение уровня белка в цереброспинальной жидкости выше 100 мг/дл, «рваные» волокна в скелетных мышцах Делеция не наследуется
2 синдрома: Синдром Пирсона –PS Гипопластическая анемия, нарушение экзокринной функции поджелудочной железы Синдром PEO– Прогрессирующая наружная офтальмоплегия Все три синдрома являются спорадическими, формиуются в зависимости от сегрегации мутантных мтДНК с накоплением в разных тканях
П.н. вместо фатального KSS может наблюдаться PEO Прогрессирующая наружная офтальмоплегия, птоз Патология связана с параличом наружных глазодвигательных мышц Процент мутантных молекул в этом случае меньше, чем при KSS синдроме, синдром не связан с угрозой для жизни больного Биохимически в мышцах обнаруживаются дефекты ферментов дыхательной цепи, особенно цитохромоксидазы
Деплеции -МDS В клетках остается 1 - 30% от нормального количества мтДНК Синдром проявляется в первые недели после рождения: фатальная гепатопатия; миопатия с генерализованной гипотонией; кардиомиопатия с судорогами (синдр. де-Тони-Дебре-Фанкони); атрофия проксимальных групп мышц; утрата сухожильных рефлексов. Смерть наступает в тяжелых случаях в первый год жизни
Генов дыхательной цепи LHON LHON+дистония Спорадическая миопатия Спорадическая миопатия Энцефаломиопатия Спорадическаямиопатия NARP MILS FBSN М Я Синдром Лея Лейкодистрофия Синдром Лея Кардиоэнцефалопатия Лейкодистрофия/тубулопатия Синдром Лея Параганглиома
Митохондриальную аномалию? При ясных симптомах – выделить кровь из вены и сделать ПЦР-анализ на точечные мутации или делеции Если результат анализа крови отрицательный, это еще не значит отсутствия заболевания (гетероплазмия!) Нужно взять биопсию: мышечную или кожную пробу у взрослых у детей Для неинвазивного тестирования используют седимент мочи, соскоб внутренней поверхности щеки, реже волосяные фолликулы
Митохондриальную аномалию? (2) Свежую мышцу анализируют гистологически и гистохимически Проводятся измерения активности отдельных звеньев комплексов дыхательной цепи «Рваные» мышечные волокна выявляются при окраске на сукцинатдегидрогеназную активность или с помощью Гомори “trichrome stain” культура фибробластов свежая мышца Если обнаруживается дефект в одном звене, это указывает на мутацию соответствующей субъединицы (я или м), если дефекты множественные – возможен дефект мт тРНК либо ядерных генов, участвующих в работе митохондрий
Митохондриальную аномалию? (3) Иногда дефект проявляется при нагрузке (NARP синдром при мутации гена ATPase6) –нужно клиническое тестирование: физические нагрузки с замерами лактата, магнитно-резонансной или инфракрасной спектроскопией Наконец, в случае еще не описанных, редких «private» мутаций проводят прямое секвенирование мтДНК
Заболеваний вовлеченность разных органов и одновременное проявление внешне не связанных между собой аномалий Наружная офтальмоплегия с нарушением проводимости сердечной мышцы и мозжечковой атаксией Мигрени с мышечной слабостью Энцефало- миопатия с диабетом Тошнота, рвота с оптической атрофией и кардиомиопатией Диабет с глухотой Глухота с наружной офтальмоплегией, птозом и ретинопатией Низкорослость с миопатией и инсультоподобными эпизодами Экзокринная дисфункция поджелудочной железы с сидеробластной анемией Задержка развития или потеря навыков и офтальмоплегия, офтальмопарез
Митохондриальные болезни? Частота митохондриальных энцефалопатий определяется примерно как 1: 11.000 Общая частота митохондриальных заболеваний – как 1: 8.000 Возраст манифестациимитохондриальных заболеваний сильно варьирует ~ 50 % после 5 лет ~ 50% - до 5 лет Смертность от митохондриальных заболеваний составляет 5-20% в год от даты манифестации
Митохондриопатия, то после перенесенных инфекционных заболеваний его состояние может резко ухудшиться также отягощают состояние стресс, голодание, переохлаждение, продолжительная обездвиженность, прием седативных средств Осторожно применять местную и общую анестезию!
Болезней –насколько это реально? Фармакологический подход Витамины, кофакторы, «ловцы» свободных радикалов – для предотвращения повреждения дыхательной цепи Наиболее успешный пример – дихлорацетат, применяемый для уменьшения лактоацидоза у пациентов с МELAS Успех частичный и временный, чаще терапия неэффективна
Болезней (2) Другой подход - уменьшить соотношение мутантная:нормальная мтДНК I. Увеличить количество немутантных молекулпутем «сдвига генов» Обычно сателлитные клетки пролиферируют и сливаются со скелетными миофибриллами в ответ на стресс или упражнение У некоторых больных с миопатией % мутантной мтДНК в сателлитных клетках ниже, чем в в скелетной мышце Пропорция нормальных мтДНК молекул в мышце увеличивалась, дефект корректировался Индуцируется пролиферация сателлитных клеток в скелетных мышцах
Болезней (3) II.Уменьшить количество мутантных молекул мтДНК Разработка синтетических молекул, избирательно связывающихся с мутаными ДНК и блокирующих их репликацию Введение в митохондрии фермента рестриктазы, избирательно разрушающего мутантную ДНК Успех достигнут пока только in vitro
Болезней (4) «Молекулярно-внутриклеточная реконструкция» Импорт из цитоплазмы нормальных тРНК вместо дефектных митохондриальных Замена дефектного комплекса дых. цепи на нормальный, полученный из другого организма (дрожжей) Пересадка ядра яйцеклетки из мутантной цитоплазмы в нормальную Все эти подходы - в стадии экпериментальной разработки
Болезней –насколько это реально? Вылечить от митохондриального заболевания сегодня невозможно Применяется симптоматическое лечение: Физическое Физиотерапия, аэробная гимнастика, умеренные и легкие нагрузки Анти-эпилептические препараты, гормоны, витамины, метаболиты, кофакторы Фармакологическое Блефаропластика, имплантация cohlear, трансплантация сердца, почек, печени, подкожная эндоскопическая гастротомия, cricopharyngeal миотомия Хирургическое
Митохондриальные заболевания или отягощает их течение Вальпроат: увеличивает частоту судорог при MELAS, гепатотоксичен Аспирин, фенобарбитал Кортикостероиды Тетрациклин, хлорамфеникол Аминогликозидыстрептомицин, гентамицин, амикацин, неомицин, канамицин - ототоксичны Этамбутол (провоцирует проявление LHON) Статин (провоцирует проявление MELAS) Антиретровирусные препараты: AZT – zidovudine, doxorubicin вызывают деплецию мтДНК Список далеко не полный!
Load More ...